|
Pollination Ecology of Lizard Orchid (Himantoglossum caprinum) in Crimea S. P. Ivanov1, A. V. Fateryga1,2, V. V. Kholodov1 |
 |
||||
|
Экология опыления Ремнелепестника козьего (Himantoglossum caprinum) в Крыму С. П. Иванов1, А. В. Фатерыга1,2, В. В. Холодов1 |
|||
|
1 - Таврический национальный университет им. В. И. Вернадского, Украина, spi2006@list.ru 2 - Никитский ботанический сад – Национальный научный центр, Украина, fater_84@list.ru |
|||
|
Abstract Приводятся данные по составу опылителей, уровню посещения цветков и опыления безнектарной орхидеи ремнелепестника козьего (Himantoglossum caprinum). Уровень опыления низкий, колеблется в разные годы и в различных пунктах Крыма от 1% до 10%, но обычно составляет 5–6 %. Как посетители цветков, способные извлекать поллинарии, зарегистрированы 9 видов пчел из семейств Megachilidae (7 видов) и Apidae (2 вида). Эффективными опылителями являются пчелы Megachile ericetorum (в основном, самцы). Анализируется уровень посещения и опыления цветков в зависимости от плотности пчел и их кормовых растений. Обсуждается способ привлечения опылителей на цветки. |
|||
|
Введение
Himantoglossum caprinum (M. Bieb.) K. Koch – редкая безнектарная наземная орхидея, описанная из Крыма (Флора СССР, 1935). По современным данным ареал вида охватывает Крым и Кавказ (Голубев, 1996). В Крыму H. caprinum распространен в горной части полуострова (в основном, в предгорьях) и на склонах южного берега. Основные места произрастания вида – светлые леса из можжевельника и дуба, лесостепи и степные склоны. Как правило, H. caprinum произрастает одиночно или группами до нескольких десятков экземпляров (Лукс, 1978; Косых, Голубев, 1983). Только в одном из известных местообитаний вид произрастает в относительном изобилии – на остепненных и покрытых кустарниками склонах Лисьей бухты в окрестностях Карадагского природного заповедника (восточная часть южного берега Крыма). В отдельные годы численность H. caprinum на этом участке достигала более тысячи генеративных экземпляров (Миронова, 2007). Антэкология H. caprinum изучена слабо. В Крыму ранее была известна только фенология цветения этого редкого вида (Миронова, 2007; Иванов и др., 2008). Вид обычно цветет с последней недели мая до середины второй половины июня. Также известно, что уровень опыления вида обычно низок (Іванов та ін., 2003). Никаких достоверных данных об опылителях H. caprinum ранее не было известно, за исключением указания в Красной книге Украины (2009) на то, что опылителями этого вида являются шмели. В данной статье мы представляем результаты наших исследований по составу опылителей, уровню посещения и опыления цветков этой орхидеи в различных условиях произрастания в Крыму. Материал и методы Исследования проводили в 2007, 2009 и 2010 годах на двух участках: в урочище Аян (рис. 1a) в окрестностях села Перевальное Симферопольского района (предгорный Крым) и в Лисьей бухте (рис. 1b). Некоторые дополнительные данные были получены в предыдущие годы (1992–2005) в урочище Аян и в двух пунктах произрастания в Севастопольском районе (поселок Максимова Дача и долина реки Черной). Один экземпляр пчелы с поллинарием H. caprinum, отловленный близ села Краснолесье (Симферопольский район), был обнаружен в энтомологической коллекции Таврического национального университета им В. И. Вернадского.  Рис. 1. Места произрастания, соцветия и опылители Himantoglossum caprinum: a – место произрастания в урочище Аян; b – место произрастания в Лисьей бухте; c, d – соцветия Himantoglossum caprinum в Лисьей бухте; e – самец Megachile ericetorum с поллинариями Himantoglossum caprinum, вид спереди; f – самка Megachile ericetorum с поллинариями Himantoglossum caprinum, вид сбоку. На изучаемых участках подсчитывалось число цветущих экземпляров H. caprinum, измерялась их высота, подсчитывалось число цветков в соцветиях и измерялось расстояние между цветущими растениями. Параллельно измеряли плотность цветков мелиттофильных растений на трансектах (5 параллельных трансект 1×20 м). Одновременно была посчитана и плотность пчел методом кошения энтомологическим сачком (300–900 взмахов). Также проводили индивидуальную поимку пчел на цветках их кормовых растений, чтобы обнаружить больше посетителей цветков H. caprinum с поллинариями этой орхидеи. В конце цветения просматривали все цветки на изучаемых соцветиях орхидей и проводили подсчет цветков, различая цветки четырех состояний: непосещенные цветки (поллинарий присутствует, рыльце без массул), цветки, посещенные впервые (поллинарий отсутствует, рыльце без массул), опыленные цветки (рыльце с массулами) с поллинарием и опыленные цветки без поллинария. Исходя из этого, подсчитывали уровень опыления, долю цветков, посещенных впервые, и оценивали индекс повторности посещения цветков посредством деления доли опыленных цветков на долю цветков, посещенных впервые (Иванов, Холодов, 2003). Этот индекс равен среднему числу цветков, посещаемых опылителем после посещения первого цветка. Кроме того, проводили оценку основных морфологических параметров цветка, таких как длина шпорца и ширина венчика, а также основных морфологических параметров пчел, посещающих цветки, таких как длина хоботка и ширина головы около центра наличника. Все измерения были сделаны при помощи измерительной линейки бинокуляра МБС-9. Названия растений приведены согласно чеклисту сосудистых растений Украины (Mosyakin, Fedoronchuk, 1999). Названия таксонов пчел соответствуют классификации Миченера (Michener, 2007). Описание условий цветения В Лисьей бухте ценопопуляция H. caprinum произрастала в ассоциации Elytrigietum (nodosae) festucosa (rupicolae) – teucriosum (chamaedrytis) с проективным покрытием от 60 до 90%. Число цветущих экземпляров на пробной площадке изменялось в отдельные годы от 25 до более чем 100. Высота генеративных побегов варьировала от 27 до 78 см (в среднем – 48,6±4,8, n=31, p=0,05). Число цветков в соцветии варьировало от 8 до 59 (в среднем – 27,5±2,9, n=52, p=0,05) (рис. 1c, d). На изученном участке одновременно с H. caprinum цвело 17 видов мелиттофильных растений. Среди них наиболее обильными были следующие: Teucrium chamaedrys L. (9,8–82,7 цв./м2), Inula germanica L. (1,9–17,4 цв./м2) и Bupleurum rotundifolium L. (1,7–25,2 цв./м2). Цветущие экземпляры орхидеи произрастали в этой ассоциации диффузно, в среднем на расстоянии 37,4±6,6 см (n=85, p=0,05) друг от друга. В изученном месте было зарегистрировано 32 вида пчел, принадлежащих к семействам Megachilidae (18 видов), Apidae (12 видов) и Halictidae (2 вида). В урочище Аян ценопопуляция H. caprinum произрастала в ассоциации Inulieto (asperae) – Filipenduletum (vulgaris) caricetum с проективным покрытием от 95% до 100%. Число цветущих экземпляров орхидеи в этом месте варьировало от 12 до 65. Высота генеративных побегов была почти точно такой же, как в бухте Лисья – от 29 до 80 см (в среднем – 47,6±4,2, n=29, p=0,05). Однако в то же время соцветия содержали меньшее количество цветков: число цветков на побеге варьировало от 5 до 30 (в среднем – 16,3±1,7, n=59, p=0,05). Одновременно с H. caprinum цвело 23 вида мелиттофильных растений. По обилию в ценозе наиболее массовые виды располагались следующим образом: Galium rubioides L. (155,6–480,0 цв./м2), Dorycnium herbaceum Vill. (32,9–1960,0 цв./м2) и Melilotus officinalis (L). Pall. (17,0–334,0 цв./м2). Цветущие особи H. caprinum располагались на этом участке более разрежено, чем в Лисьей бухте: среднее расстояние между экземплярами составляло 91,6±21,2 см (n=61, p=0,05). На участке было зарегистрировано 28 видов пчел. Среди них 6 видов принадлежали к семейству Megachilidae, 7 – к Apidae, 8 – к Halictidae, 4 – к Andrenidae, 2 – к Colletidae и 1 – к Melittidae. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Опылители и прикрепление поллинариев
Все экземпляры насекомых, зарегистрированные как посетители цветков H. caprinum принадлежали к длиннохоботным пчелам (табл. 1): 7 видов – к семейству Megachilidae (первые 14 экземпляров) и 2 вида – к семейству Apidae. Только пчелы Megachile ericetorum (которых было отловлено 6 экземпляров) несли по 3 или 4 поллинария на одном экземпляре (в одном случае – 2 поллинария), два экземпляра двух других видов несли по 2 поллинария, остальные 8 экземпляров остальных 6 видов несли по 1 поллинарию или висцидию, оставшемуся от поллинария. Непосредственное посещение цветков H. caprinum было замечено только в одном случае – самкой Megachile parietina (наблюдение не включено в таблицу). Пчела подлетела к соцветию и после непродолжительного зависания в воздухе быстро присела на один из цветков. После этого она вставила свой хоботок в шпорец, а затем улетела с цветка. При этом поллинарий не был извлечен, так как к этому времени его уже извлек предыдущий посетитель этого цветка.
Как правило, поллинарии H. caprinum прикреплялись к пчелам в центральной части наличника или вблизи фронтально-клипеального шва, изредка на лбу (рис. 1e, f; рис. 2c, d). Поллинарии, прикрепленные к апикальному краю наличника, зарегистрированы у Anthidium loti и у обоих экземпляров Megachile pilicrus. Чтобы извлечь поллинарии, пчела должна коснуться бурсикулы фронтальной поверхностью головы. Бурсикула прикрывает висцидии, которые располагаются чуть выше верхнего края рыльца (рис. 2a, b). Если поллинарии прикрепятся к апикальному краю наличника, массулы не смогут коснуться рыльца после реконфигурации поллинариев – их опускания, которое происходит сразу же после прикрепления поллинариев к телу пчелы. Вместо этого они коснутся внутренней полости цветка ниже рыльца – около отверстия шпорца. Таким образом, A. loti и M. pilicrus не в состоянии опылять цветки H. caprinum. Все другие виды теоретически могут опылять орхидею, но поллинарии с истраченными массулами были зарегистрированы нами только у M. ericetorum. Самцы этого вида имеют точное морфологическое соответствие цветкам H. caprinum. Их хоботок достигает 4,3–7,4 мм в длину (в среднем – 6,10±0,30 мм, n=29, p=0,05) а ширина головы – 2,3–3,9 мм (в среднем – 3,02±0,12 мм, n=29, p=0,05). В то время как у цветков орхидеи длина шпорца составляет 6,5–11,4 мм (в среднем – 9,09±0,36, n=35, p=0,05) а ширина венчика цветка – 2,4–3,7 мм (в среднем – 2,99±0,11, n=35, p=0,05). Таким образом, ширина головы у этих пчел варьирует в том же диапазоне, что и ширина венчика, а их хоботок значительно короче шпорца. В Лисьей бухте в 2007 году мы зарегистрировали несколько цветков H. caprinum с прокушенными шпорцами. Вероятно, они не были прокушены отмеченными выше посетителями цветков. Скорее всего, это было сделано шмелями Bombus argillaceus (Scopoli, 1763) или одним из трех видов пчел Xylocopa, которые часто встречались на данной территории. 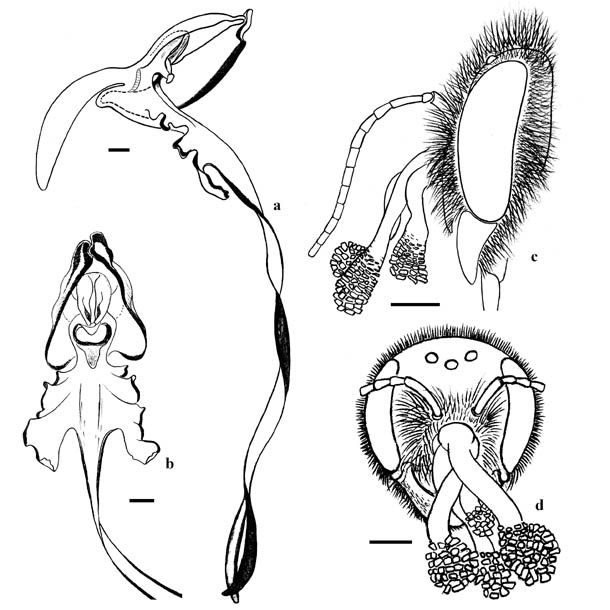 Рис. 2.Цветки Himantoglossum caprinum и местоположение поллинариев на головах пчел: а – продольный разрез цветка, вид сбоку, b – цветок, вид спереди, c – голова самца Anthidium cingulatum с поллинариями, вид сбоку , d – голова самки Megachile ericetorum с поллинариями, вид спереди. Все масштабные линейки – 1 мм.
Рис. 2.Цветки Himantoglossum caprinum и местоположение поллинариев на головах пчел: а – продольный разрез цветка, вид сбоку, b – цветок, вид спереди, c – голова самца Anthidium cingulatum с поллинариями, вид сбоку , d – голова самки Megachile ericetorum с поллинариями, вид спереди. Все масштабные линейки – 1 мм.
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Эффективность опыления Данные по уровню посещения и опыления цветков H. caprinum в условиях различной плотности пчел и плотности цветков их кормовых растений представлены в таблице 2. Доля опыленных цветков была низкой и варьировала от чуть более 2% до более чем 10%. В другие годы и на других участках мы получили сходные данные. В урочище Аян уровень опыления в другие годы составил 1,4% (в 2005) и 3,6% (в 2008). В Максимовой Даче этот показатель составил 0,6% (в 1993), 3,4% (в 1995), 5,5% (в 1992) и 6,8% (в 2000). На Черной речке уровень опыления только в одном изученном году (1993) составил 6,5%.
Доля опыленных цветков на обоих участках коррелировала с плотностью пчел, особенно длиннохоботных (табл. 2). В Лисьей бухте самый высокий уровень опыления наблюдался в 2007, когда наблюдалась самая высокая плотность длиннохоботных пчел. В этом же году была зарегистрирована самая высокая доля цветков, посещенных впервые. Высокое значение этого показателя указывает на изобилие неспециализированных посетителей цветков, которые могут извлекать поллинарии, но не могут опылять ими цветки, или которые посещают цветки только один раз (низкая повторность посещения цветков). Высокая доля цветков, посещенных впервые, также соответствует очень низкой плотности цветков кормовых растений пчел. В течение двух других годов наблюдений плотность цветков кормовых растений пчел была выше, а плотность пчел – ниже. Это обусловило более низкие показатели доли посещенных и опыленных цветков орхидеи. В урочище Аян плотность цветков была намного выше, чем в Лисьей бухте, но плотность длиннохоботных пчел была сопоставима. Таким образом, показатели доли посещенных и опыленных цветков также оказались сопоставимыми. Высокий показатель повторности посещения цветков орхидеи в урочище Аян (особенно в 2010) связан с тем, что среди длиннохоботных пчел, обитающих на этом участке, преобладал специализированный опылитель (M. ericetorum). Обсуждение Среди всех видов орхидей мелитофильный синдром опыления является наиболее широко распространенным (Черевченко та ін., 2010). Пчелы привлекаются на цветки нектаром или при помощи псевдопыльцы; имитацией обобщенного образа источника пищи (неспециализированный пищевой обман); за счет цветочной мимикрии (специализированный пищевой обман); имитацией мест ночевки; имитацией самок пчел (половой обман) или имитацией самцов пчел (псевдоантагонизм). Неспециализированный пищевой обман является среди мелитофильных видов орхидей наиболее распространенным. У цветков этих видов нет явного сходства с цветками определенных вознаграждающих симпатрических видов растений. Орхидеи, использующие обобщенную имитацию пищи, получают эффективное опыление тогда, когда они цветут раздельно от вознаграждающих нектаром видов растений. Это достигается фенологическим разделением между орхидеями и вознаграждающими видами: первые цветут обычно раньше последних (Иванов и др., 2008; Pellissier et al., 2010). У H. caprinum нет близкого сходства с цветками кормовых видов растений (Иванов и др., 2003), в урочище Аян этот вид начинает цвести спустя месяц после цветения видов, использующих обобщенную имитацию источника пищи (например, Orchis simia Lam.) и цветет при более высокой плотности цветков кормовых видов. У этой орхидеи также наблюдается самый низкий уровень опыления, в то время как у других шести видов орхидей с подобным механизмом привлечения он составляет приблизительно 35–60% (Иванов и др., 2008). Таким образом, H. caprinum приспособился к существованию при низком уровне опыления, что обеспечивается некоторыми специфическими механизмами привлечения опылителей. Пчелы семейства Megachilidae редко отмечаются в качестве эффективных опылителей безнектарных орхидей. В Европе представители этого семейства (олиголектные виды рода Chelostoma) зарегистрированы как эффективные опылители только у пыльцеголовника красного (Cephalanthera rubra (L). Rich.), цветки которого имеют мимикрическое сходство с цветками кормовых растений этих пчел из рода Campanula (Nilsson, 1983; Назаров, Иванов, 1990). H. caprinum является вторым видом орхидей, который опыляется пчелами-мегахилидами, и он опыляется , как и в случае с C. rubra, одним или несколькими близкородственными видами пчел. Преобладание самцов среди посетителей цветков H. caprinum позволяет предположить, что этому виду орхидей свойственен половой обман. Однако присутствие нескольких самок M. ericetorum среди опылителей, которые несли на себе поллинарии с истраченными массулами, и наблюдения за поведением самки M. parietina свидетельствуют о наличии пищевого обмана. Этот обман является неспециализированным (без определенной цветочной мимикрии), так как посетители цветков орхидеи были, главным образом, полилектными пчелами. Конкретные факторы, которые привлекают на цветки главным образом мегахилид (особенно M. ericetorum), а не другие виды пчел, неизвестны. Вероятно, это может быть специальный олфакторный аттрактант, который эффективен только для мегахилид. Эта гипотеза не лишена оснований, поскольку у другого близкородственного вида орхидей из рода Steveniella обнаружен аттрактант, который эффективен только для складчатокрылых ос, особенно подсемейства Vespinae (Nazarov, 1995). Специализированные олфакторные аттрактанты для ос-веспид также зарегистрированы и у вознаграждающего вида орхидей – дремлика зимовникового (Epipactis helleborine (L). Crantz) (Brodmann et al., 2008). Среди не орхидей другое вознаграждающее растение – норичник теневой (Scrophularia umbrosa Dumort.) привлекает на свои цветки различных ос-веспид, и при этом некоторые виды (главным образом их самцы) особо эффективно привлекаются и даже становятся "пьяными" после многих повторных посещений (Фатерыга и др., 2006). Самки и самцы M. ericetorum также демонстрируют постоянно повторяемые посещения цветков H. caprinum, все они несли несколько поллинариев, большинство из которых были с истраченными массулами. Случай с самцом, несущим три поллинария без истраченных массул, означает, что он посетил три цветка за такой короткий промежуток времени, что извлеченные поллинарии еще не успели опуститься. Таким образом, H. caprinum приспособлен к опылению пчелами и, наиболее вероятно, его цветки испускают специальный |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Литература
Голубев В. Н. Биологическая флора Крыма / Издание второе. – Ялта: НБС – ННЦ, 1996. 126 с. Иванов С. П., Фатерыга А. В., Тягнирядно В. В. Сравнительная оценка эффективности опыления орхидей в урочище Аян // Бюлл. Гос. Никитск. ботан. сада. 2008. Вып. 97. С. 10-14. Иванов С. П., Холодов В. В. Анализ характера опыления безнектарных орхидей (Orchidaceae) в зависимости от их пространственного размещения // Вопросы развития Крыма. Симферополь, 2003. Вып. 15. С. 57-65. Іванов С. П., Кобечинська В. Г., Отуріна І. П., Пилипенко Н. В. Динаміка цвiтiння та ефективність запилення безнектарних та нектарних видiв орхiдей в Криму // Питання бiоiндикацiї та екології. 2003. Вип. 8, № 2. С. 43-50. Косых В. М., Голубев В. Н. Современное состояние популяций редких, исчезающих и эндемичных растений горного Крыма. Ялта, 1983. 119 с. Лукс Ю. А. Классификация орхидей флоры Крыма по категориям редкости // Бюлл. Гос. Никитск. ботан. сада. 1978. Вып. 3. С. 15-18. Миронова Л. П. Редкие виды // Летопись природы (Карадагского природного заповедника). Симферополь, 2007. С. 87-140. Назаров В. В., Иванов С. П. Участие пчел рода Chelostoma Latr. (Hymenoptera, Megachilidae) в опылении мимикрирующих видов Cephalanthera rubra (Z.) Rich. и Campanula taurica Juz. в Крыму // Энтомол. обозр. 1990. Т. 69, № 3. С. 534-537. Фатерыга А. В., Иванов С. П., Новиков Е. В. Складчатокрылые осы (Hymenoptera: Vespidae) – специализированные опылители редкого вида норичников – Scrophularia umbrosa (Scrophulariales: Scrophulariaceae) в Крыму // Изв. Харьковск. энтомол. об-ва. 2006. Т. 14, вып. 1-2. С. 145-161. Флора СССР / Ред. В. Л. Комаров. Ленинград, 1935. Т. 4. 760 с. Червона книга України. Рослинний світ / Ред. Я. П. Дідух. Київ: Глобалконсалтинг, 2009. 912 с. Черевченко Т. М., Буюн Л. І., Ковальська Л. А. Запилювальні стратегії в орхідних (Orchidaceae) // Укр. ботан. журн. 2010. Т. 67, № 5. С. 637-649. Brodmann J., Twele R., Francke W., Ayasse M. Pollinator-attracting semiochemicals of the wasp-flower Epipactis helleborine // Mitt. Deutsch. Ges. Allg. Ang. Entomol. 2008. Bd. 16. S. 171-174. Michener C. D. The bees of the World / 2nd ed. Baltimore, 2007. xvi + 953 p. Mosyakin S. L., Fedoronchuk M. M. Vascular plants of Ukraine: a nomenclature checklist. Kiev, 1999. xxiii + 345 p. Nazarov V. V. Pollination of Steveniella satyrioides (Orchidaceae) by wasps (Hymenoptera, Vespoidea) in the Crimea // Lindleyana. 1995. Vol. 10. № 2. P. 109-114. Nilsson L. A. Mimesis of bellflower (Campanula) by the red helleborine orchid Cephalanthera rubra // Nature. 1983. Vol. 305. № 5937. P. 799-800. Pellissier L, Vittoz P., Internicola A. I., Gigord L. D. B. Generalized food-deceptive orchid species flower earlier and occur at lower altitudes than rewarding ones // J. Plant Ecol. 2010. Vol. 3. № 4. P. 243-250. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
republished 5.10.2011 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Published in: I. I. Shamrov (ed.). Okhrana i kultivirovaniye orkhidey [Protection and cultivation of orchids] (Materials of the 9th International conference, St.-Petersburg, September 26–30, 2011). – Moscow: KMK Scientific Press Ltd., 2011. – P. 187–194.
Опубликовано в: Охрана и культивирование орхидей (Материалы IX Международной конференции, Санкт-Петербург, 26–30 сентября 2011 г.). – Москва: Товарищество научных изданий КМК, 2011. – С. 187–194. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
© 2011 by Authors |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||